
By Vidja Rajan, Columnist, The Times
The previous group of Inner Nature articles were loosely organized around how organisms sense the external environment using vision, sound, touch, taste and smell. What is most remarkable about this cluster of what appears to be completely different mechanisms is the profound underlying similarity. This similarity is due to the outcome of notionally different sensory similar mechanisms that similarly trigger nerve impulses to the brain where they are interpreted.
Trial and error by the mechanism scientists call evolution has, over millenia, developed the ability of organisms to link attractive stimuli and aversive stimuli to behavioral outcomes in progressively more complex ways. We can follow the stepwise increase in complexity of these senses by examining the spectrum from single-celled organisms to progressively more complex organisms. But because this is an insanely difficult thing to do for each sense, I have, in my previous Inner Nature articles on various senses, examined only two groups of organisms: complex mammals such as ourselves and evolutionarily less complex invertebrate animals such as insects.
In the next set of articles, starting with this one, I will examine physiological functions that underpin multicellular organisms. As with my previous articles, I will compare vertebrate mammals with invertebrate insects. This first article will be on food, because it is the source of energy which underlies life. Without energy, there is no life. Food is just energy stored in chemical bonds, and the process of digestion releases that energy and makes it available for life processes to take place.
Biomolecules are organic, a term used by chemists to describe molecules that contain carbon. Carbon is joined to other atoms using chemical bonds. Joining atoms together to make molecules requires energy and breaking chemical bonds releases energy. Plants, for example, use photosynthesis to harness energy from the sun to join the atoms obtained from carbon dioxide and water, and which they then use to build their bodies. Those biomolecules can feed animals, who eat plants and sometimes each other, and break them down in their digestive systems.
The main groups of dietary biomolecules for animals are proteins, carbohydrates and fats. They have slightly different compositions and are joined by slightly different types of bonds. Therefore, the bonds have slightly different shapes and energies. For example, fat bonds store 9 Calories per gram whereas a gram of carbohydrates packs only 4 Calories. Bonds from different dietary molecules also have different shapes. To release these stored calories, those bonds have to be broken by enzymes, but, due to their different shapes, they require different tools. Just as a spanner does not do the same job as a hammer, enzymes specialize: proteins are disassembled by enzymes called proteases, the carbohydrate starch by amylases, and fats by lipases. There is more fine tuning than that – just as there is not just one size of spanner or hammer, there are many types of proteases for breaking down proteins. Not to belabor the point, but this is true for all enzymes – they are very specific for their substrates – they behave like keys to unlock specific locks.
It’s these enzyme tools that operate to disassemble specific bonds in different organs of the digestive system. Food enters through the mouth, passes through the stomach, and then makes its way through the small and large intestines, and undigested waste is expelled through the anus. Carbohydrate-digesting amylase enzymes are present in saliva. The stomach efficiently digests proteins. The first part of the small intestine, the short duodenum, does the bulk of the work of enzymatic digestion with a variety of proteases, fat-digesting lipases, a slew of carbohydrate-digesting amylases, lactases and maltases. Subsequently, digested food is absorbed into the blood by the microvilli of the next two longish portions of the small intestine, the jejunum and ileum. Blood carrying digested food then passes through the liver where detoxification of ingested poisons like alcohol occurs. The remaining digested food passes through the large intestine, where water is absorbed, and finally expelled from the body. Beneficial microorganisms populate the entire digestive canal and provide a variety of functions including digesting and absorbing nutrients, and immune regulation [1].
Digestive systems are adapted to foods eaten. Mammals can be grouped into omnivores, carnivores and herbivores. The digestive systems of omnivores like humans are largely like carnivores (sorry, vegans!) We have shortish guts which digest proteins and fats effectively but are unable to digest cellulose, a carbohydrate, which passes through the system as fiber. Herbivores, on the other hand, have longer digestive systems with regions populated by symbiotic cellulose-digesting single-celled organisms called protists. But that’s not all. There are actually two groups of herbivores – ruminants/foregut herbivores like cows which have a neutral to slightly alkaline rumen where cellulose-digesting protists reside anterior to the acidic stomach. The food they swallow is regurgitated and chewed again as “cud” before passing through the reticulum into the omasum where fats are absorbed. It is only then that food enters the true stomach (abomasum) before it passes on to the small intestine. These animals slowly but efficiently digest their food because chewing the cud and swallowing the much-masticated gloop causes fiber to be broken down thoroughly. Then there are the hindgut herbivores, such as horses and rabbits, which have a single acidic stomach through which the grasses pass before entering the small intestine. A large diverticulum called the cecum extends from the junction of the small and large intestine where symbiotic protists that digest cellulose reside. This means that food is digested again in the cecum, but these animals do not extract a lot of benefit from this step because the large intestine is not as efficient with absorption. Hindgut herbivores have three choices to get more benefit from the food based on their lifestyle – consuming their still-nutritive droppings to process it a second time (rabbits and hares), graze constantly to keep ingesting nutrients (horses), or get really, really, large so as to eat a lot at one sitting to obtain enough nutrients to keep going (elephants). Figure 1 summarizes these digestive systems.
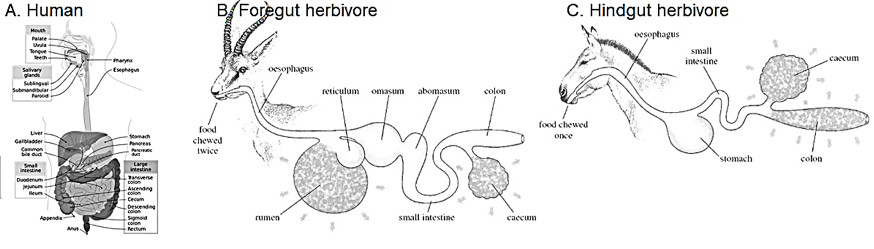
Figure 1: Comparison of human, foregut and hindgut digestive systems. Humans have monogastric digestion typical of a carnivorous gut. Foregut herbivores have multiple chambers, anterior to the acidic abomasum, housing microbiota typical of ruminants. Horses have a hindgut digestion with a large cecum that houses cellulose-digesting microbiota posterior to the acidic stomach.Credits: Figure A from By Mariana Ruiz, Jmarchn – Own work, Public Domain, https://commons.wikimedia.org/w/index.php?curid=2898771and Figures B and C from https://www.open.edu/openlearn/ocw/mod/oucontent/view.php?printable=1&id=2220&extra=thumbnail_idm36655888 CC-BY-NC-SA-4.0
Arthropods, of which insects are a class, are one of the most abundant animal phyla on the planet and were the first animals to colonize land. Because of all the habitats and food sources available to them, they diversified enormously. They also have a bewildering variety of diets as a consequence, from grass to other animals, to each other. For example, parasitic insects such as female Strepsipterans have lost almost all their body parts, reverting to a larval grub-like stage, absorbing nutrients from the host’s hemocoel [2]. In free-living insects, mouthparts protrude from the four anterior segments, the labrum, mandible, maxilla, and labium. The labrum is an extension of the head and contains chemoreceptors on the periphery. The main cutting and chewing apparatus in most insects is the mandible, which has two hinged parts which meet in the middle with teeth-like cusps that fit into each other, slicing food with hard, sclerotized edges. The maxilla are loaded with chemosensors and have hard, pointed ends which can be used to manipulate food for further cutting before it is swallowed. Sucking insects like bees have mandibles and maxilla modified to form a tube sheathed by the labium (see Figure 2A-D)
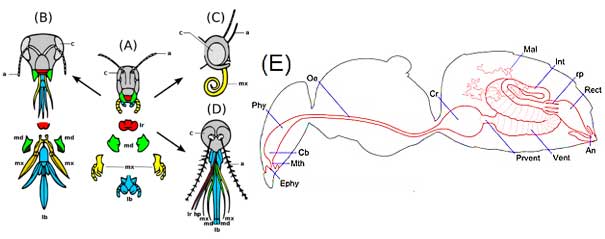
Figure 2: The development of insect mouthparts from the primitive chewing mouthparts of a grasshopper in the centre (A), to the lapping type (B) of a bee, the siphoning type (C) of a butterfly and the sucking type (D) of a female mosquito. (E) Bee gut schematic: foregut is in yellow, midgut in orange and hindgut in green. Legend: a, antennae; c, compound eye; lb, labium; lr, labrum; md, mandibles; mx, maxillae hp hypopharynx. An – anus, Cb – cibarium, Cr – crop, Ephy – epipharynx, Int – intestine, Mal – Malpighian tubules, Mth – mouth, Oe – oesophagus, Phy – pharynx, Prvent – proventriculus, Rect – rectum, rp – rectal pad, Vent – ventriculusA-D from: Xavier Vázquezderivative work: Siga – Evolution insect mouthparts coloured.png, CC BY-SA 3.0 https://commons.wikimedia.org/w/index.php?curid=15923055; Bee gut schematic (E) from http://honeybee.drawwing.org/book/digestive-system
An insect’s digestive system is divided into three parts: the foregut, midgut and hindgut [3](Figure 2E). The foregut contains the buccal cavity with optional salivary glands, the pharynx, esophagus and crop. The pharynx is surrounded by muscles which can crush food, and the crop is often a storage organ for food. In bees, the crop stores nectar which is later regurgitated in the hive to make honey. Passing into the midgut, the actual digestive region requires food to pass through a gate called the proventriculus. Once in the midgut, food is digested by a variety of enzymes. The midgut contains anterior tubular extensions called gastric cecae, blind tubes which may increase digestive surface area. The hindgut is separated from the midgut by a sphincter which regulates passage of food. The junction between midgut and hindgut is embellished with many hair-like extensions called Malpighian tubules, the insect equivalent of kidneys. The hindgut functions in absorption of digested nutrients. Fecal material is mixed with nitrogenous waste supplied by the Malpighian tubules and excreted outside the body.
Finally, and significantly, many insects and some amphibians undergo complete metamorphosis, changing their diets as they change their morphology and much of their physiology. Insects change from a grub-like form to a winged form; frogs develop from aquatic, herbivorous tadpoles to terrestrial, carnivorous forms. This is significant, because larvicides such as Bacillus thuringiensis (Bt) are proteins which are only toxic to animals with alkaline gut contents, such as caterpillars. But foregut herbivores also have neutral to alkaline guts. Acidic stomachs digest proteins, so larvicidal proteins such as Bt do not affect monogastric and hindgut herbivores. While there has not been much research on this subject, my inference is that insect larvae, aquatic tadpoles, and foregut herbivores such as cows, will be more susceptible to Bt GMOs than carnivores, omnivores, hindgut herbivores (horses), or even adult frogs and adult insects. Any research scientists who want to test this hypothesis out there?
- Stevens, C.E. and I.D. Hume, Contributions of microbes in vertebrate gastrointestinal tract to production and conservation of nutrients. Physiological reviews, 1998. 78(2): p. 393-427.
- Mazumdar, A., Twisted winged endoparasitoids. Resonance, 2004. 9(10): p. 19-24.
- Terra, W.R. and C. Ferreira, Insect digestive enzymes: properties, compartmentalization and function. Comparative Biochemistry and Physiology Part B: Comparative Biochemistry, 1994. 109(1): p. 1-62.




